
Elasmobranchii teeth and Teleostei otoliths
Universidad Francisco de Miranda, Centro de Investigaciones Arqueológicas,
Antropológicas y Paleontológicas (CIAAP), Calle Zamora, Balcón
de Los Arcayas,
Zona Colonial, Coro, Edo. Falcón. Venezuela.
aguilero@unefm.edu.ve,
dione19@hotmail.com
Synopsis of Neogene Caribbean Fishes *
In an effort to document the extent and distribution of the Neogene
marine fishes in time and space, we are analyzing samples of the Caribbean
Neogene sedimentary environments along the Costa Rica, Panama, Venezuela
and Trinidad Basins (Aguilera et. al., man.). We have identified
altogether ~ 205 taxa, based in 50,000 teleostean otoliths and 2,000 elasmobranch
teeth in 265 collections from 26 formations from the South Caribbean Sea
(Fig. 1), ranging in age from approximately 19.0 to 1.0 million years.
This synopsis represents a progress report on our ongoing work to consistently
identify and describe the taxa. It includes diagnostic characters of the
fish otoliths and teeth from different sedimentary basins and ages.
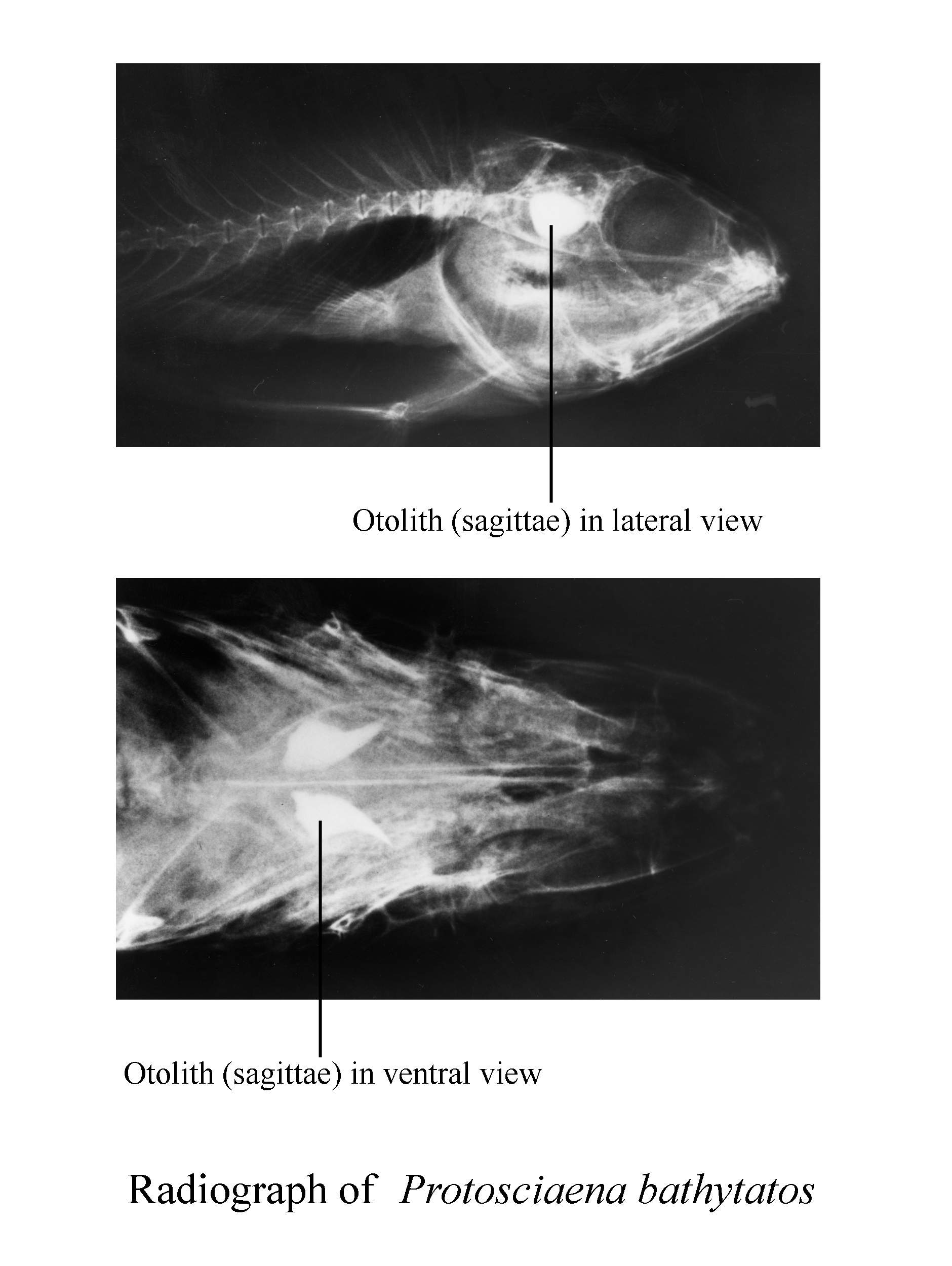
Figure 2. Location of otoliths within a fish [Sciaena bathytatos
(Perciform:Sciaenidae)]
Otoliths are individual elements in the complex skeletal architecture
in the teleostean fishes (Figs. 2, 3). They have been discussed extensively
by Nolf (1985) including their anatomy, morphology, variability, ontogenetic
changes, and preservation. As he noted, otoliths are the integral, specialized
hard part of the actinopterygian and sarcopterygian acoustico-lateralis
system, situated in the membranous labyrinths. Each labyrinth is located
on either side of the brain in the otic capsules of the neurocranium. Within
each membranous labyrinth, there are different otoliths, in the utriculus,
in the sacculus and in the lagena. When the term "otolith" is used in this
study, it refers to the saccular otolith except in catfish, which are represented
by utricular otoliths. Otoliths consist of calcium carbonate, mainly in
the form of aragonite, and organic matter called otoline.

Figure 3. General outline of the morphology of an otolith showing
diagnostic features
Otoliths are useful in the definition of characters in phylogenetic analyses of fish, e.g., the phylogeny of the Sciaenids (Sasaki, 1989). However, the exclusive use of otoliths in phylogenetic analyses of taxa such as the Sciaenids does not produce clearly resolved relationships among taxa (Schwarzhans, 1978). Moreover, controversial descriptions of new fossil taxa and the frequent use of taxa in open nomenclature further compromise their utility in taxonomic analyses and have produced much confusion in paleobiogeographic analyses.
Excellent preservation and abundant ontogenetic series of fossil otoliths are uncommon throughout most Caribbean Neogene sedimentary basins. In addition, an enormous effort has been required to make extensive collections of large ontogenetic series of comparative Recent material, which represent ~25,000 valid living species (Eschmeyer et.al., 1996). The ideal conditions suggested by Nolf (1985), therefore, do not exist for making adequate taxonomic counts. Exponential increases in the numbers of new taxa based on fossil otoliths (Huyghebaert and Nolf, 1979) provide only general estimates, and a decade of work is still needed to obtain a more realistic picture of the diversity of Neogene teleostean otoliths. The same is true in living fishes, of which ~200 300 new species are still being described each year (Eschmeyer et.al., 1996).
Detailed reviews of the fossil record of Caribbean Neogene teleostean
otoliths have been published by Nolf (1976), Nolf
and Stringer (1990), Nolf and Aguilera (1998) and Stringer (1998).
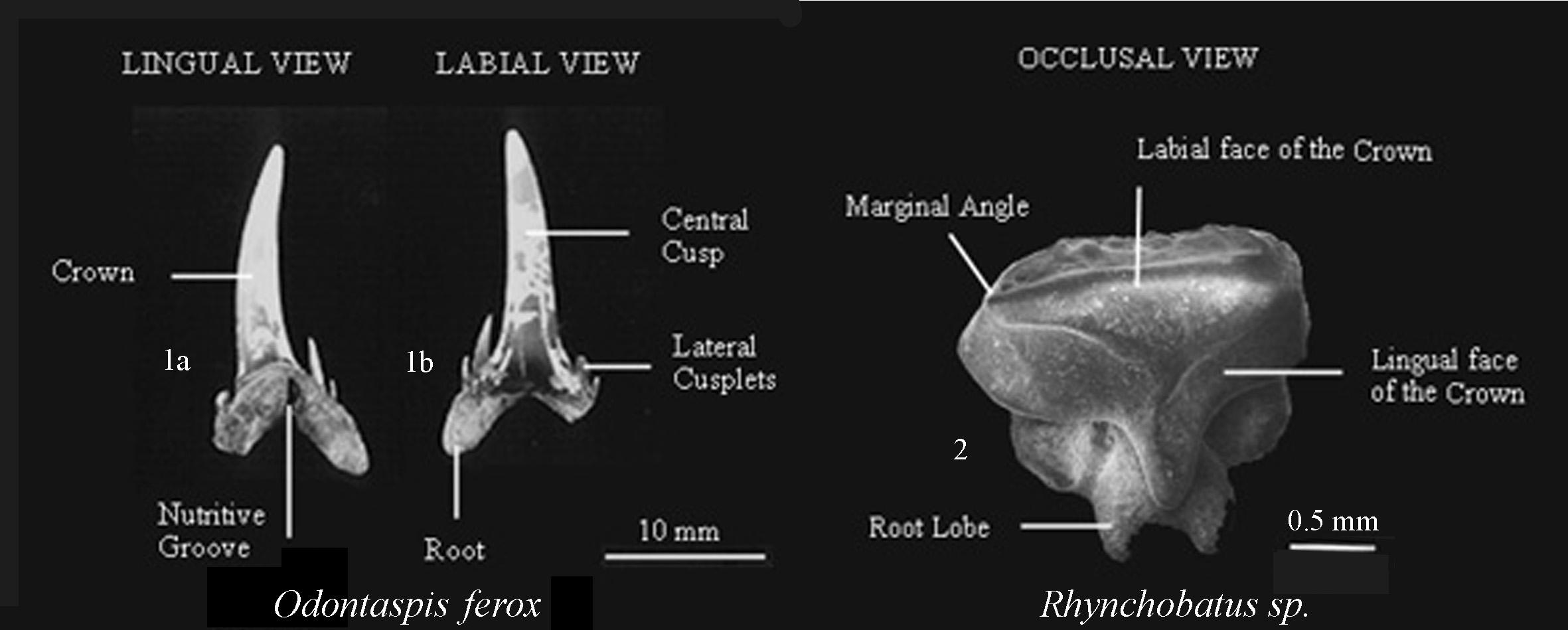
Figure 4. General outline of the morphology of shark teeth and a
rays tooth showing diagnostic features
Oral teeth (Fig. 4), rostral teeth, dental plates, dermal denticles
and caudal spines represent the more common
anatomical fossil elements used in the systematics of the Elasmobranchii
(Cappetta, 1987). Heterodonty and sexual dimorphism need to be considered
in such work. Large dental series of fossil specimens and extensive collections
of comparative Recent selachian species are also required.
Unfortunately, numerous undescribed living species of sharks and stingrays exist in the world today. An example is the Australian selachians (Last and Stevens, 1994). Systematic reviews of Caribbean Neogene elasmobranchs have not been attempted since Leriche (1938, 1942), Casier (1958), Gillette (1984) and Iturralde-Vincent et.al. (1996). However, a recent compilation by Kruckow and Thies (1990) provides information attesting to the scarce and disperse records of fossil selachians in South Caribbean sedimentary basins.
According to Aguilera et.al. (manuscript), these previous records
represent a only small fraction in the richness of the Neogene Caribbean
fish fauna (Fig. 5), because the numbers of taxa collected closely parallel
sampling effort, suggesting that a large fraction of taxa from each age
interval is uncollected. This is confirmed by the steep increase in
cumulative numbers of taxa as a function of the numbers of collections
or specimens, whether broken down by age and basin,
or for all collections combined (Fig.6).
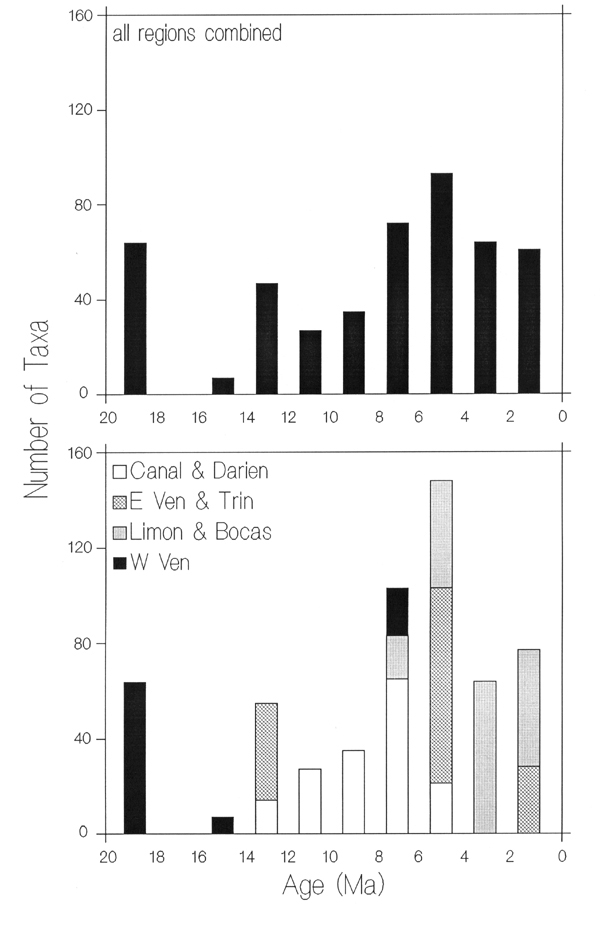
Figure 5. Known richness of Neogene Caribbean fish species.
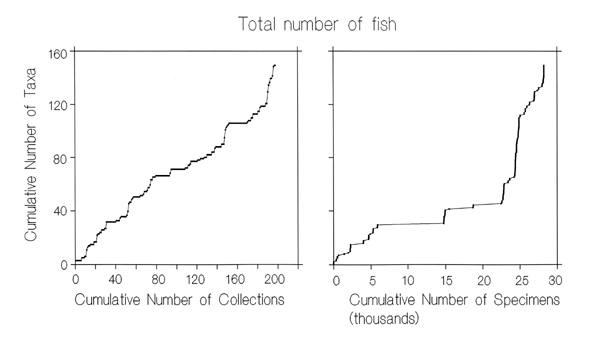
Figure 6. Sampling of Neogene Caribbean fish.
The reasons for these daunting results are that most collections contain comparatively few specimens or taxa and, as expected in the tropics (Sanders 1969; Jackson et.al., 1999), most taxa are extremely rare.
The classification of fishes was arranged in part following Eschmeyer et.al. (1996), with the inclusion of fossil taxa following Cappetta (1987) and Nolf (1985). Diagnostic characters were established using Smale et al. (1995). Formal descriptions of new taxa and undetermined species are still in progress. Otoliths and teeth micrographs were made with a scanning electron microscope using a retro dispersed electron technique.
*Panama Paleontology Project, Smithsonian Tropical Research Institute
